Francis Martin
Laboratoire d’excellence ARBRE, UMR Interactions Arbres/Micro-organismes, Centre INRAE Grand Est-Nancy

Les arbres forestiers remplissent leurs fonctions d’absorption de l’eau et des éléments minéraux du sol grâce à une interaction symbiotique mutualiste entre leurs racines et une grande diversité de champignons. La sylviculture et la trufficulture mettent déjà ces connaissances en pratique, et les progrès de la recherche en biologie et en écologie dans ce domaine des interactions plantes/micro-organismes ouvrent de nouvelles perspectives d’application à la gestion durable et à la protection des forêts face aux changements climatiques.
Malgré les menaces multiples qui pèsent sur la plupart des environnements naturels, bois, forêts et prairies hébergent encore une richesse mycologique insoupçonnée. Des milliers d’espèces de champignons peuplent les forêts. A l’automne, cèpes, chanterelles, rosés des prés, coprins et lactaires sont récoltés en abondance et les fricassées de ces produits de la terre agrémentent de nombreuses tables familiales. Dans le langage populaire, ce qu’on désigne couramment comme «champignon» et que l’on cueille en forêt et dans les prairies n’est en fait que l’appareil reproducteur, une fructification temporaire et visible appelée le sporophore («qui porte les spores») par les mycologues. Le plus souvent composé d’un pied et d’un chapeau protégeant lamelles ou tubes, il est la production d’un organisme souterrain à caractère plus durable et plus discret constitué d’un lacis de filaments ouateux formés d’hyphes, le mycélium végétatif. Ce dernier émerge d’une spore sous forme d’un tube germinatif et se ramifie rapidement dans le sol, la litière et dans pratiquement tous les organes des arbres vivants ou morts. Le mycélium constitue la partie principale du champignon et il peut former des colonies pouvant couvrir plusieurs dizaines de mètres carrés et représenter jusqu’à 99% du poids de l’organisme. En dehors des sporophores, les champignons peuvent aussi être observés sous forme de spores, de rhizomorphes (faisceaux de plusieurs filaments mycéliens agrégés en fibres), de mycorhizes avec les racines des arbres ou de structures d’infection chez de nombreuses plantes.
On reconnaît trois grands groupes de champignons correspondant à différents modes de nutrition, en particulier vis-à-vis de l’acquisition du carbone :
- les décomposeurs, ou saprotrophes, qui dégradent et consomment efficacement les polysaccharides et les protéines de la nécromasse végétale, microbienne et animale,
- les symbiotes, qui s’alimentent des sucres simples (glucose) provenant de leur plante-hôte, mais en ayant des effets bénéfiques pour leur partenaire. Leurs réseaux mycéliens souterrains explorent le sol et la litière à la recherche d’éléments minéraux qu’ils transportent et transfèrent à leur plante-hôte en échange des sucres importés,
- les parasites, ou pathogènes, qui infectent les plantes et les animaux et détournent à leur profit les sucres et les acides aminés solubles accumulés dans les tissus colonisés, entraînant l’affaiblissement et parfois la mort de leur(s) hôte(s).
Les champignons décomposeurs jouent un rôle crucial en assurant la productivité et la stabilité à long terme de tous les écosystèmes terrestres, dont les forêts. En effet, ils assurent la libération des éléments minéraux de la nécromasse (par exemple, les feuilles et le bois mort) et leur recyclage dans la biomasse, permettant ainsi le maintien de la fertilité et la pérennité de la production primaire, essentiellement assurée par les arbres. Les champignons appartenant aux deux autres catégories, souvent regroupées sous le terme de biotrophes, sont plus directement responsables à court ou moyen terme de l’état du peuplement puisqu’ils vivent en partie à l’intérieur des tissus des arbres et affectent positivement (symbiotes mutualistes) ou négativement (parasites) leur métabolisme, leur physiologie et leur croissance. Les champignons symbiotiques ectomycorhiziens représentent très souvent plus de 50% de la communauté fongique d’une forêt dans les régions tempérées ou boréales.
Les mycorhizes des arbres forestiers
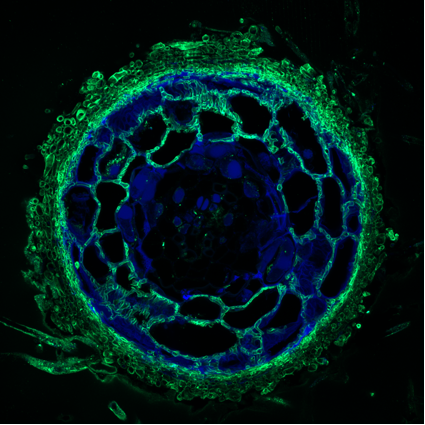
Dans nos forêts caducifoliées ou de résineux, les racines des grandes essences sylvicoles (par exemple Pins, Epicéa, Chênes, Hêtre) portent généralement des ectomycorhizes qui peuvent être formées par plusieurs centaines d’espèces de champignons basidiomycètes (par exemple Amanites et Bolets) et ascomycètes (par exemple, les Truffes). Morphologiquement, on distingue une racine courte ectomycorhizée d’une racine courte non mycorhizée par la présence du manteau fongique externe recouvrant la racine mycorhizée (Fig. 1). Les filaments mycéliens de ce manteau se développent d’une part vers l’intérieur de la racine, en s’insinuant entre les cellules de l’épiderme et du parenchyme cortical racinaire pour former le réseau de Hartig et d’autre part, un réseau mycélien qui explore le sol. La croissance du réseau ramifié de filaments mycéliens externes peut être considérable (jusqu’à 1000 m de mycélium par mètre de racine), augmentant ainsi considérablement le volume de sol exploité par l’arbre mycorhizé. Le réseau mycélien qui explore ce volume de sol est capable de mobiliser des éléments minéraux (par exemple le phosphate inorganique) inaccessibles aux racines de la plante-hôte et de les transférer, sous leur forme initiale ou après les avoir transformés, à leur associé végétal. Outre les ressources minérales solubles (phosphate, azote minéral), les champignons ectomycorhiziens sont également capables d’altérer les minéraux et d’exploiter une partie des ressources accumulées dans la matière organique des sols en sécrétant des protéases, des phosphatases ou des polyphénols oxydases. On comprend facilement que l’établissement de ces associations symbiotiques ectomycorhiziennes apparaisse comme une stratégie importante pour assurer la survie et la croissance des arbres.
(A)

(B)
Mais comment le champignon symbiotique reconnaît-il son partenaire végétal ? Comment la plante-hôte distingue-t-elle le champignon bénéfique du champignon parasite ? Comment le champignon évite-t-il les défenses immunitaires de la plante ? Tout est question de dialogue biochimique et moléculaire ! Les racines de la plante apte à contracter la symbiose libèrent dans le milieu qui les entoure des quantités infinitésimales de molécules «signales». La perception de ces signaux chimiques déclenche la libération par le champignon de petites protéines dans les racines qu’il colonise. Celles-ci se lient au récepteur de l’acide jasmonique, l’hormone de défense de la plante, neutralisant ainsi les réactions défensives qu’aurait dû induire la présence massive du champignon dans la racine. Le champignon symbiotique contrôle ainsi la réponse immunitaire de la plante colonisée. Il peut alors s’établir et développer un troc avec la plante «sous influence» qui l’héberge.
Pendant plus d’un siècle, l’étude de la symbiose mycorhizienne a reposé sur l’idée simpliste d’une association bénéfique entre une plante et un champignon uniquement. Les recherches actuelles conduites sur le terrain révèlent cependant que ces associations mutualistes établissent des interactions multiples impliquant plusieurs partenaires. La symbiose mycorhizienne structure un véritable réseau de plantes et de champignons d’espèces différentes, reliés entre eux : jusqu’à plusieurs centaines d’espèces de champignons par arbre et une vingtaine d’arbres colonisés par un même champignon. Des plantes voisines, même d’espèces différentes, peuvent donc partager des champignons communs. L’étude des implications fonctionnelles et écologiques de ces réseaux tissés entre champignons et arbres débute et nécessite la mise en place d’expérimentations complexes sur le terrain.
Les réseaux mycorhiziens établissent des flux nutritifs entre plantes
Lors d’une expérience, menée dans les années quatre-vingt-dix par le professeur Suzanne Simard, de jeunes bouleaux et sapins de Douglas ectomycorhizés par le même champignon, ont été marqué par l’apport de CO2 enrichi en isotopes 13C et 14C. Ces marquages révélèrent que les arbres, connectés par les réseaux mycorhiziens souterrains, recevaient chacun du carbone, vraisemblablement des sucres solubles, l’un de l’autre, avec un flux net en faveur du sapin de Douglas. La quantité de sucres reçus par ce dernier représentait de 10 à 25% des produits de sa photosynthèse. Le réseau ectomycorhizien est donc bien impliqué dans les transferts entre bouleau et sapin de Douglas. Une expérience plus récente, réalisée dans une forêt du Jura suisse, a permis d’évaluer qu’environ 4% des composés carbonés issus de la photosynthèse d’un arbre sont transportés vers les arbres voisins connectés au même réseau ectomycorhizien. Ces mesures instantanées du flux de carbone ne permettent néanmoins pas de calculer l’impact global sur la plante durant toute la saison de végétation et sur son budget nutritionnel à long terme.
Le réseau mycorhizien permet l’échange d’autres ressources que les sucres entre plantes, comme le démontrent des dispositifs simples où on cultive deux plantes mycorhizées dans un même sol. On a ainsi pu démontrer des flux d’azote, de phosphate, ou encore d’eau par le réseau symbiotique.
La communication entre les partenaires symbiotiques
Terminons en mentionnant l’un des effets les plus surprenants du réseau mycorhizien sur les plantes qu’il connecte : la mise en place d’un dialogue impliquant des signaux d’alerte. Dans une plante infectée par un champignon parasite, ou grignotée par une chenille, des réactions de défense se mettent en place qui limitent l’attaque de l’agent infectieux et qui impliquent la production d’hormones de stress, comme l’éthylène ou le jasmonate de méthyle. Ces composés volatils déclenchent une cascade de réactions de défense aboutissant à l’accumulation de molécules toxiques pour l’intrus, par exemple de puissants inhibiteurs des enzymes digestives de l’insecte phytophage. Dans des expérimentations réalisées en chambre de culture, sous conditions contrôlées, des plantes proches reliées par un réseau mycorhizien souterrain, quant à elles vierges de toute attaque, mettent en place des défenses similaires quelques jours après l’attaque de leur voisine ! On ignore encore la nature des signaux (hormones, dépolarisations membranaires) et la façon dont ils transitent via les hyphes dans les conditions naturelles de la forêt, mais il n’y a pas de doute que les plantes dialoguent via leur réseaux mycéliens. Arbres et champignons forment ainsi d’immenses réseaux sociaux qui irriguent forêts, bois et prairies. L’activité de ces réseaux interconnectés est cruciale afin d’assurer le fonctionnement durable des écosystèmes, et leur maîtrise pourrait bien aider les forestiers à atténuer les effets désastreux du changement climatique.